


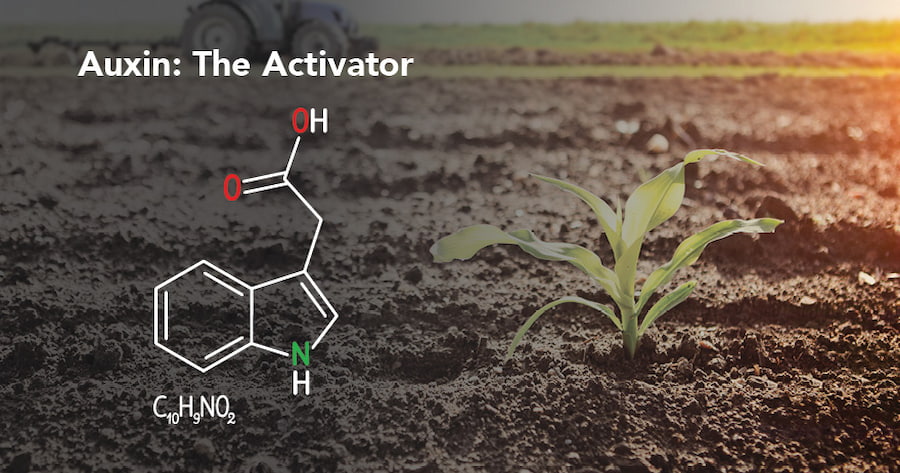
اکسین تنظیم کننده طبیعی رشد گیاه
اکسین یا ایندول – ایل – استیک اسید (IAA) نوعی تنظیم کننده طبیعی رشد گیاه درون زاد بوده. نقش اساسی در تقسیم تمایز و طویل شدگی سلولهای گیاهی دارد. در سطح اندام و گیاه کامل اکسین تاثیری عمیق روی بسیاری جنبه های فیزیولوژی گیاه شامل مورفولوژی گیاهچه، زمین گرایی، نور گرایی، غالبیت انتهایی، پیری و ریزش برگ، گلدهی میوه دهی و رسیدگی میوه دارد.
اکسین تنظیم کننده طبیعی رشد گیاه
اکسین در بافتهای مریستمی فعال در حال رشد و از مسیرهای وابسته به تریپتوفان و یا مستقل از تریپتوفان ساخته شده است. در سرتاسر پیکره گیاه در غلظتهای مختلف بین ۱ تا ۱۰۰ میکروگرم IAA در هر کیلوگرم از وزن تر گیاه یافت میشود. غلظت اکسین در گیاهچه های جوان و بافتهای در حال رشد و طویل شدگی سریع بیشتر از بافتهای بالغ است. بنابراین میتوان گفت بافتهای جوانتر به این تنظیم کننده رشد حساسیت بیشتری دارند. در طی ۶۰ سال گذشته بررسیهای زیادی روی اثرات اکسین های بیرونی بر رشد گیاه صورت گرفته و نتایجی بدست آمده که عبارتند از :
- اکسینها بسته به غلظت موجود میتوانند محرک یا بازدارنده رشد گیاه باشند.
- بافتهای مختلف حساسیت متفاوتی به مصرف اکسین دارند.
چرا از اکسین ها در علف کش ها استفاده می شود؟
ممانعت از رشد در غلظتهای بالاتر از حد نرمال اکسین عمدتاً مربوط به اتیلن تولیدی القاء شده توسط اکسین است. بنابراین وقتی مقدار اکسین در یک بافت خاص به یک سطح بحرانی میرسد، اتیلن تولید میگردد و به طور نسبی مانع رشد میشود. کلائوس گروسمن و همکاران (۲۰۰۱) در تحقیقات خود در شرکت BASF روی علف هرز بی تیراخ تیمار شده با علفکشهای اکسینی نشان دادند که بین تخریب بافت و تولید پراکسیدهیدروژن (H2O2) ارتباط است. آنها متوجه شدند که یکی از پیامدهای حاصله از تیمار علف هرز با علفکشهای اکسینی تحریک سنتز اتیلن و افزایش بیوسنتز هورمون آبسیزیک اسید (ABA) است.
غلظت اکسین در یک بافت زنده به شدت تحت کنترل سرعت نسبی بیوسنتز و تجزیه ترکیبات الحاق شده است. از سال ۱۹۴۷ به بعد مشخص شد که بافتهای گیاهی قادر به تجزیه اکسیداتیو IAA هستند. این تجزیه توسط آنزیم IAA اکسیداز صورت گرفته که فعالیت سریع و گسترده ای در بافتهای گیاهی دارد. اگرچه فعالیت این آنزیم کاملاً مشخص نیست، اما به طور قطع میتوان آن را از پراکسیدازهای گیاه جدا کرد. غلظت اکسین در بافتهای در حال طویل شدن در نتیجه فعالیت کم آنزیم اکسیداز نسبتاً بالا (تقریباً ۱۰۶ مولار)، و در ریشه در نتیجه فعالیت زیاد آنزیم اکسیداز پائین تقریباً ۱۰-۱۰ مولار است.
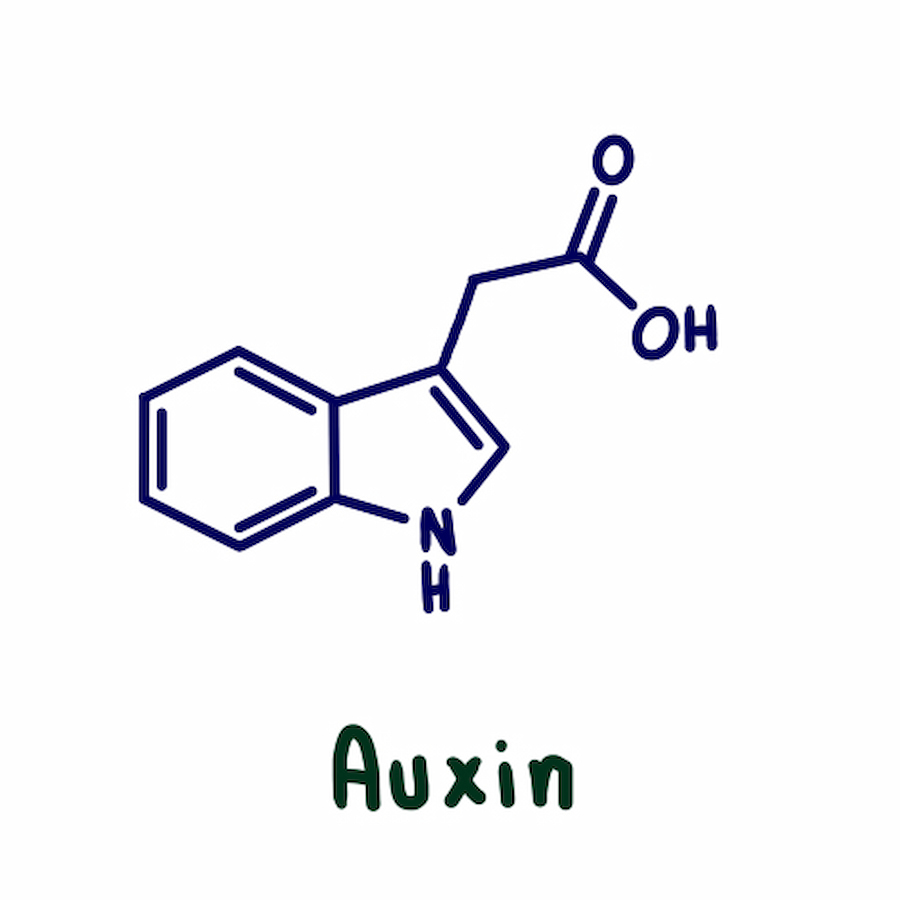
هورمون اکسین در گیاهان
اکسین با الحاق به گلوکز اسیدهای آمینه و میواینوسیتول ۲ میتواند در گیاه ذخیره شده و در زمان نیاز به ویژه پس از جوانه زنی در پی هیدرولیز به اکسین آزاد تبدیل شود. بررسیهای اخیر نشان میدهد اهمیت و اثر فیزیولوژیکی غلظتهای اکسینهای الحاقی در محیط بافت زنده بیشتر از IAA آزاد است. به نظر میرسد که در بافتهای رویشی، آسپارتات اسید آمینه اصلی الحاق شده به اکسین بوده که به فرم -IAA آسپارتات یافت میشود.
اکسین
این الحاق توسط آنزیم ال آسپارتات-ان-آکیلاز آنزیم القائی تمام اکسینهای طبیعی و سنتز شده صورت میگیرد. استرهای گلوکز نیز از ترکیبات الحاقی معمول با اکسین هستند که از گلوکوزیل ترانسفرازهای از قبل موجود تشکیل شده اند. هرچند کار انجام نشده در این رابطه زیاد است، ولی به نظر میرسد برهمکنش بین سنتز اکسین تجزیه و الحاق آن تعیین کننده غلظت اکسین طبیعی در گیاه است.
گیرنده های اکسین؛ تظاهر اثر ژن و علفکشها
قبل از اینکه اکسین متابولیسم سلول و رشد بافت را تغییر دهد، باید ابتدا به گیرنده خود متصل شده و علامتی به دستگاه متابولیکی سلول ارسال نماید. یک گیرنده اکسین را شاید بتوان به عنوان جایگاه مولکولی دقیقی از سلول تعریف کرد که طی یکسری واکنش باعث تغییراتی در رشد میشود. بنابراین مکانیسم اولیه عمل اکسین، اتصال به گیرنده اکسین است. وینز (۱۹۸۵) پیشبینی نمود این اتصال باید:
- برگشت پذیر باشد، چراکه وقتی اکسین حذف میشود واکنشها کند یا متوقف میشوند.
- میل ترکیبی بالائی داشته باشد. چراکه غلظت اکسین های درونزاد گیاه بسیار کم است.
- قابل اشباع باشد. در غلظتهایی مشابه با غلظتهایی که فرآیندهای فیزیولوژیکی حساس به اکسین اشباع میشوند.
- اختصاصی باشد تا فقط با اکسین ها فعال شود.
- محدود باشد به بافتهای حساس به اکسین محدود باشد.
- به یک پاسخ بیولوژیکی مربوط شود.
تحقیق در مورد گیرنده های هورمونی گیاه بسیار کمتر از تحقیقات مربوط به سیستمهای باکتریای و حیوانی است در سالهای اخیر علی رغم پیشرفتهای سریع علمی مسائل مبهم زیادی باقی است. دو گیرنده اصلی برای اتصال به اکسین مطرح است:
- پروتئین ABP1 (پروتئین پیوند اکسینی)
- پروتئین TIR1 (پروتئین پاسخ بازدارنده انتقال
انتقال اکسین در گیاه
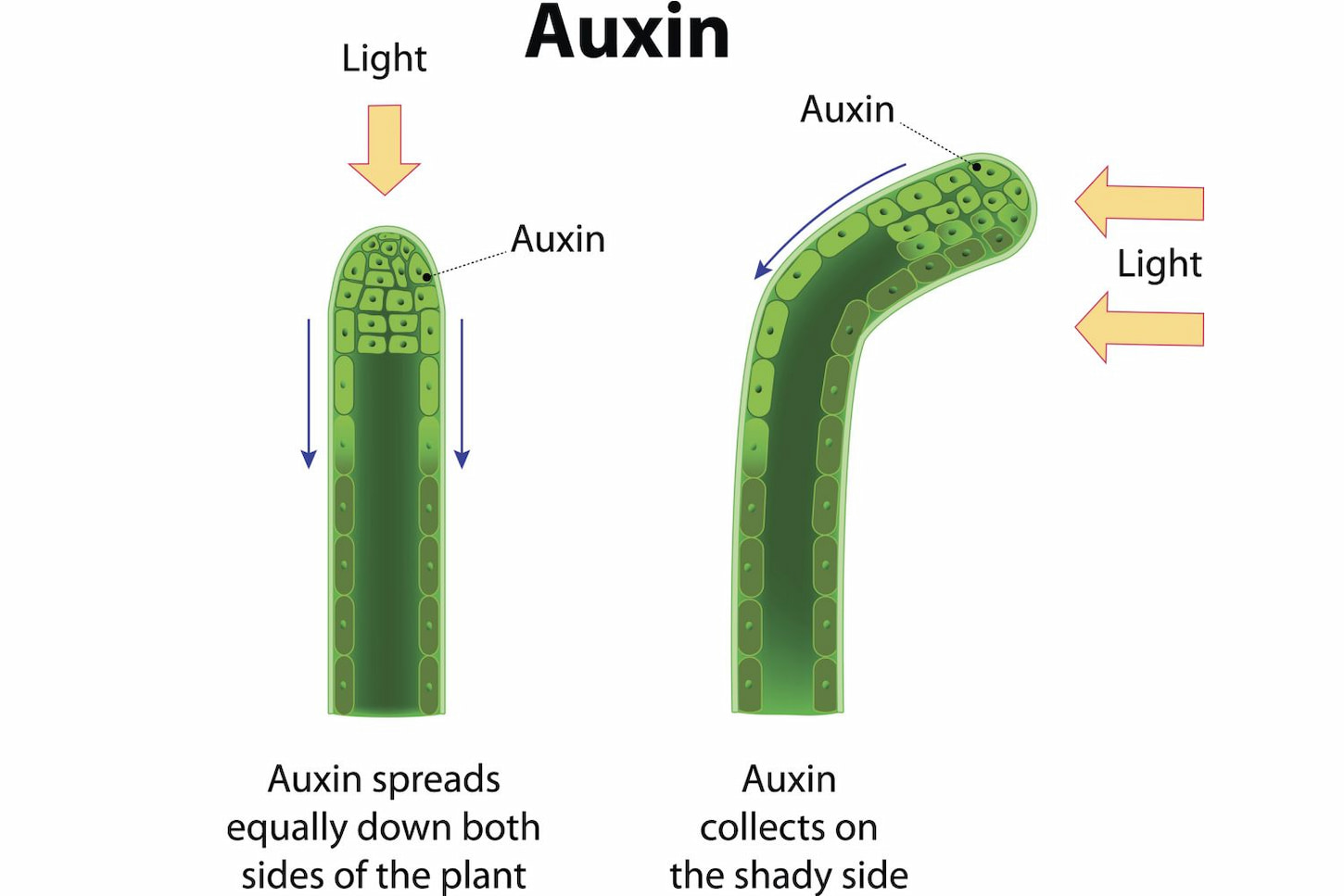
اکسین تنظیم کننده طیف وسیعی از فرایندهای تکاملی گیاه است. این تنظیم کنندگی ناشی از توزیع نامتقارن آن در سلول و بافتهای همجوار در مراحل مهم رشد و توسعه گیاه است. اکسین در بافتهای مریستمی سنتز و از طریق آوند آبکش در سرتاسر گیاه توزیع شده یا از طریق سیستم انتقال قطبی به صورت کنترل شده از یک سلول به سلول دیگر منتقل میشود.

فرضیه شیمیواسمز، مدل جدیداً پذیرفته شده برای انتقال قطبی اکسین است. مولکول IAA ابتدا در دیواره سلولی به شکل پروتون دار IAAH حضور داشته و به صورت غیر فعال از غشای پلاسمائی عبور میکند. پس از ورود به داخل سلول پروتون جدا شده و مولکول به صورت یون IAA در داخل سلول به دام می افتد. برای انتقال فرم یونی IAA به بیرون سلول، به حاملهای مخصوص انتشار اکسین نیاز است. این حاملهای به طور نامتقارن در سلول قرار گرفته، باعث انتقال یکطرفه اکسین میشوند. امروز مشخص شده حاملین اکسین قادر به جذب IAA نیز هستند.
انتقال اکسین در گیاه
ماهیت حامل انتشار اکسین با مطالعه PIN1 در موتانت شاهی گوش موشی مشخص شد. این گیاه موتانت دارای ساقه های سوزن مانند فاقد گل بود. تجزیه مولکولی ژن PIN1 در سال ۱۹۹۸نشان داد که این ژن یک پروتئین حامل برای عبور از غشا را کد میکند. بعدها ۷ ژن PIN دیگر نیز شناسایی شدند که فنوتیپ حاصل از کلیه این ژنها با تجمع غیر عادی اکسین مرتبط بود. پروتئینهای PIN طبق فرضیه شیمیو اسمز به روش قطبی قرار می گیرند.
چگونه حاملهای اکسین میتوانند به جایگاه های مشخصی در سلول منتقل شوند؟
ویتن و همکاران (۲۰۰۷) در بررسیهای خود جابجایی و مقصد حاملها را مسیر تردد ریزکیسه های وزیکولهای موجود در سیتوپلاسم پیشنهاد کردند. تصور بر این است که پروتئین کمکی AXR4 تضمین کننده قرارگیری پروتئین در غشای پلاسمائی است، اما چگونگی این کار هنوز مشخص نشده است. در شاهی گوش موشی پروتئینهای PIN از طریق اندوزومها به سرعت بین غشای پلاسمائی و شبکه آندوپلاسمی در چرخش هستند. این فرآیند که چرخش ساختاری خوانده می شود طبق نظر پچیورک و همکاران (۲۰۰۵) توسط اکسین ممانعت میشود.
این چرخش ساختاری در کنار اکسین هایی که از نظر بیولوژیکی فعال هستند، باعث افزایش حاملهای انتشار اکسین در سطح سلولی جایی که غلظت اکسین حداقل است، میشود. از این رو اکسین ها میتوانند میزان انتشار خود را افزایش دهند. اغلب تصور بر این است که ممانعت از انتقال اکسین میتواند مکانسیم موثری برای بهره برداری از علفکش باشد.
-
دیفلوفنزوپیر در ترکیب با علفکش اکسینی مانع رشد علف های هرز پهن برگ
ترکیبی که در سالهای اخیر با این ویژگیها معرفی شده، دیفلوفنزوپیر (۶۶۲ BAS) است. دیفلوفنزوپیر به تنهایی مانع رشد علف هرز شده، اما در ترکیب با یک علفکش اکسینی باعث افزایش انتقال علفکش اکسینی به نقاط انتهایی علف هرز و کنترل موثرتر علف هرز پهن برگ میشود. ترکیب دایکامبا با دیفلوفنزوپیر باعث افزایش طیف کنترل علفهای هرز پهن برگ و تحمل ذرت در دزهای نسبتاً پائین ۱۰۰ تا ۳۰۰ گرم ماده موثره در هکتار میشود.
دیفلوفنزوپیر با اتصال به پروتئین خاص، از انتقال اکسین به مریستمهای انتهایی ممانعت میکند. این علفکش با ۱۵۰ معادل ۱۹ میل ترکیبی بالایی به این محل دارد. بنابراین تجمع اکسین های طبیعی و یا سنتز شده در نقاط انتهایی گیاه القاء کننده پاسخ به دز بیش از حد اکسین است. دیفلوفنزوپیر همین طور مانع زمین گرایی ریشه میشود.








دیدگاهتان را بنویسید